بحث پیدایش و تکامل تولید مثل جنسی، به عنوان یکی از گذار های تکاملی عظیم، بسیار مورد توجه است. پیچیدگی تولید مثل جنسی از یک سو و تنوع آن از سوی دیگر، به رازآلود بودن این مسئله می افزایند.
به ادامه مطلب مراجعه کنید.
شکل زیر، اولین گام را از سناریویی ممکن برای منشا جنسیت نشان می دهد. نخستین مرحله جانداری تک سلولی را با چرخه زندگی هاپلوئید - دیپلوئید نشان می دهد. در بخشی از چرخه، سلول ها به صورت هاپلوئید رشد می کنند و در بخشی دیگر به صورت دیپلوئید. پس جمعیت بین دو وضعیت هاپلوئید و دیپلوئید در تناوب است. از این نظر، این نوع چرخه زندگی مشابه چرخه زندگی برخی جانداران مدرن دارای تولید مثل جنسی است؛ البته شیوه تحول از مرحله هاپلوئیدی به دیپلوئیدی و مرحله دیپلوئیدی به هاپلوئیدی در این دو نوع از جانداران متفاوت است.

مرحله اول
سلول هاپلوئید در این نوع چرخه برای دیپلوئید شدن، با سلول هاپلوئید دیگر لقاح نمی کند بلکه به نحوی ساده، با تکثیر هر کروموزوم و بدون تقسیم شدن، دو مجموعه کروموزوم به دست می آورند. این فرایند ساده با عنوان "میتوز درونی (endomitosis)" شناخته شده است که امروزه در برخی سلول ها رخ می دهد. سلول دیپلوئید نیز با شکلی بسیار ساده و تک مرحله ای از میوز، به دو سلول هاپلوئید تقسیم می شود.
از آن جا که موضوع منشا میوز است و به نظر می رسد توضیح آن سخت باشد، لحظه ای بحث را از مسئله منحرف می کنیم و به آن بر می گردیم. پیش از وقوع میوز، هر سلول دو نسخه از هر کروموزوم را حمل می کند. پس از اتمام میوز، هر سلول یک نسخه از هر نوع کروموزوم را حمل می کند. حین میوز هر کروموزوم با همتای خود که "همولوگ (homologue)" نامیده می شود، جفت شده و سپس از هم جدا شده و به سمت یکی از قطبین دوک حرکت می کند. میوز کارآمد به جفت شدن صحیح همولوگ ها بستگی دارد. جالب است بدانیم در اغلب موارد عقیم یا نازا بودن دورگه ها، به علت تفاوت بیش از اندازه کروموزوم های والدی است که مانع از جفت شدن آن ها می شود.
در نخستین گامی که توصیف شد، یک "میوز تک مرحله ای" را تصور کردیم که در آن خبری از مضاعف شدن کروموزوم ها و کراسینگ اور [مانند آن چه در پروفاز I امروزی رخ می دهد] نیست. این ها پیچیدگی هایی هستند که بعد ها مورد بحث قرار می گیرند. تمام آن چه رخ می دهد این است که هر کروموزوم با همتای خود جفت شده، از یکدیگر جدا شده و سلول تقسیم می شود. جایی برای کراسینگ اور وجود ندارد؛ دو کروموزوم همولوگ در این نوع چرخه زیست، کاملا یکسان هستند چرا که تنها چند نسل قبل میوز، از راه میتوز درونی تولید شده اند.
چنین نوعی از میتوز تک مرحله ای، صرفا یک فرضیه نیست و امروزه در برخی آغازیان تک سلولی رخ می دهد اما در حال حاضر نمی دانیم آیا کراسینگ اور در چنین آغازیانی رخ می دهد یا خیر و برای دانستن آن نیازمند مطالعات ژنتیکی بیشتر هستیم.
چرخه هاپلوئید- دیپلوئید یاد شده، به عنوان نیای میوز پیشنهاد شده و کمابیش پذیرفتنی است. هم میتوز درونی و هم میوز تک مرحله ای در جانداران کنونی یافت شده اند. اما چه مزیت انتخابی وجود داشته است که منجر به پیدایش دیپلوئیدی شده است؟ احتمالا دیپلوئیدی پاسخی به محیط متغیر، شاید چرخه تغییرات سالانه، بوده است. دلایل خوبی وجود دارد که چرا دیپلوئیدی در برخی محیط ها یک مزیت است. DNA آسیب دیده تنها در صورتی می تواند ترمیم شود که مولکولی سالم برای نسخه برداری وجود داشته باشد. این امر نیازمند دیپلوئیدی است. آسیب DNA در غلظت بالای اکسیژن معمول است. بنابراین احتمالا دیپلوئیدی نوعی سازش در برابر دوره های افزایش غلظت اکسیژن بوده است. و اما مزیت مربوط به هاپلوئیدی کدام است؟ [به هر حال، بخشی از چرخه یاد شده هاپلوئیدی را در بر می گیرد]. سلول های هاپلوئید کوچک تر هستند و بنابراین، نسبت سطح به حجم بیشتری دارند. به همین دلیل، در کمبود غلظت مواد غذایی مخمر های هاپلوئید سریع تر از انواع دیپلوئید رشد می کنند.
اما مانعی سر راه توضیح یاد شده وجود دارد. سلول های مخمر با انتشار مولکول ها از سطح خود تغدیه می کنند. اما می دانیم نخستین یوکاریوت ها با فروبردن ذرات تغذیه می کرده اند. بنابراین با مسئله ای مواجه شده ایم که امید است پاسخ آن را در مطالعه روی آغازیان کنونی بیابیم.
نخستین مرحله ای که توصیف شد ارتباطی با جنسیت ندارد. فایده پیشنهاد چنین مرحله ای کمک به توضیح منشا شکل ساده ای از میوز است. در دومین مرحله، که در شکل زیر مشاهده می شود، میتوز درونی با لقاح جایگزین شده است. این مرحله، یک چرخه جنسی واقعی است.

مرحله دوم
دو دلیل می توانیم برای ایجاد این تغییر پیشنهاد کنیم: آشکار ترین دلیل این است که لقاح می تواند آثار جهش های مضر مغلوب را پوشش دهد. بین دو سلول هاپلوئیدی متفاوت، یکی از آن ها می تواند حامل جهش باشد. اگر جهش مغلوب باشد، سلول دیپلوئید حاصل از لقاح، از تاثیرات منفی جهش در امان خواهد بود. این همان مکانیسمی است که تحت عنوان پدیده "قدرت دورگه (heterosis)" خوانده می شود: فرد حاصل از دورگه گیری دو سویه درون آمیخته [1] نسبت به والدین خود، توان و قدرت بیشتری دارد. اما مزیت لقاح نسبت به میتوز درونی می تواند اساسی باشد.
دلیل محتمل دوم به این شرح است که امکان دارد که یک ترانسپوزون خودخواه برای کسب اطمینان از تکثیر شدن خود، پدیده لقاح را رقم زده باشد. بیشتر ژن ها تنها هنگامی همانند سازی می کنند که سلول تکثیر شود. اما عناصری متحرک یا اصطلاحا ترانسپوزون هایی وجود دارند که به مکان جدیدی در طول ژنوم پرش می کنند در حالی نسخه ابتدایی همچنان سر جای خود وجود دارد. بنابراین پس از نقل مکان (transposition) دو نسخه از ترانسپوزون وجود دارد؛ در حالی که پیش از نقل مکان تنها یک نسخه از آن وجود داشت. فرض کنید این اتفاق در جانداری با تولید مثل جنسی رخ دهد. تعداد نسخه های ترانسپوزون می توانند افزایش یابند اما تعداد جاندارانی که نسخه های ترانسپوزون در آن حضور دارند افزایش نمی یابد. بقای طولانی مدت این نسخه ها تماما به تکثیر میزبان آن ها وابسته است.
اما فرض کنید چنین ترانسپوزونی میزبان خود را وادار به لقاح با سلولی دیگر کند؛ در آن صورت قادر است به کروموزوم هایی که از سلول دیگر آمده اند نقل مکان کند. در چنین حالتی اگر سلول تقسیم شود ترانسپوزون مذکور در هر سلول دختر موجود خواهد بود در حالی که پیش از این تنها در یک والد حضور داشت. به عبارت دیگر، با وادار کردن سلول به لقاح، ترانسپوزون راهی دیگر برای تکثیر خود یافته است. چنین عنصر ژنتیکی می تواند در یک جمعیت گسترش یابد حتی اگر این امر همان طور که انتظار می رود، تا حدی شایستگی میزبان خود را کاهش دهد.
دونال هیکی (Donal Hickey) و مایکل رز (Michael Rose) در سال 1988 پیشنهاد دادند که لقاح جنسی نه به دلیل فایده ای برای سلول ها بلکه به علت فایده ای که برای تکثیر ترانسپوزون ها داشته است، آغاز شده است.
این ایده به دو دلیل جذابیت دارد. اولا بی درنگ مزیت انتخابی خاصی را برای لقاح جنسی ایجاد می کند: دیگر نیازی به جستجوی مزیت بلند مدت لقاح برای جمعیت نداریم. در ثانی، پیشینه ای از چنین عناصر ژنتیکی در باکتری ها وجود دارد. پلازمید ها، عناصر ژنتیکی فرعی هستند، معمولا حامل تعداد اندکی ژن هستند و در اغلب باکتری ها حضور دارند.
این پلازمید ها نمی توانند سلول های میزبان خود را که واجد دیواره ای سخت هستند وادار به لقاح کنند اما برخی پلازمید ها قادرند سبب اتصال باکتری میزبان خود به باکتری دیگر شوند. با این اتصال یک نسخه از DNA پلازمید به باکتری دیگر منتقل می شود. گاهی حتی بخشی از DNA کروموزوم باکتریایی می تواند به سلول دریافت کننده منتقل شود [2].
سناریوی پیشنهادی هیکی و رز چندان بعید نیست. اگر حق با آن ها باشد، به نتیجه ای نسبتا متناقض می رسیم: لقاح جنسی از این رو پیدایش یافته است که به سود یک ترانسپوزون بوده است در حالی که برای سلول میزبان این فرایند هزینه بر بوده است؛ اما در دراز مدت، جنسیت به علت فایده ای که برای جمعیت داشته، باقی مانده است.
در سومین گام از نظریه پیشنهادی که در شکل زیر قابل مشاهده است، میوز تک مرحله ای ساده و بدون کراسینگ اور با میوز دو مرحله ای همراه نوترکیبی، جایگزین شده است. اولین سوالی که پیش می آید این است: نوترکیبی (recombination) چه مزایایی دارد؟ یک جمعیت دارای تولید مثل جنسی دارای دو مزیت نسبت به جمعیت های فاقد تولید مثل جنسی است: سریع تر تکامل می یابد و کمتر در معرض خطر جهش های مضر قرار می گیرد.

مرحله سوم
حال جمعیتی را با تولید مثل جنسی، لقاح و میوز در نظر بگیرید و فرض کنید نرخ نوترکیبی ژن ها در افراد این جمعیت متفاوت است. این فرض قابل قبول است: در تمام جمعیت های دارای تولید مثل جنسی که با دقت مطالعه شده اند، ژن ها دارای نرخ نوترکیبی متفاوتی بوده اند. همان مزیت های تولید مثل جنسی برای جمعیت، برای ژن ها نیز صادق است. اگر محیط سریعا متغیر باشد یا جهش های مضر به صورت همزمان رخ دهند، انتخاب طبیعی ژن های دارای نرخ نوترکیبی بالا را ترجیح خواهد داد.
دومین سوالی که پاسخ به آن سخت تر است، این است: چرا در میوز کنونی ابتدا کروموزوم ها همانند سازی می کنند و سپس دو تقسیم سلولی رخ می دهد تا هاپلوئیدی حاصل شود؟ چرا میوز باید این دو مرحله را شامل شود؟
در توصیف هر تکست بوک پایه از میوز، ابتدا هر کروموزوم مضاعف می شود و چهار نسخه از هر نوع کروموزوم وجود خواهد داشت. سپس کراسینگ اور بین رشته های همولوگ رخ می دهد و در پی آن با دو بار تقسیم سلولی، گامت های هاپلوئید تولید می شوند. اما این کتب ندرتا درباره شگفتی حقیقی این فرایند اظهار نظر می کنند. کاربرد اولیه میوز، نصف کردن عدد کروموزومی است: پس چرا باید با دو برابر شدن این عدد آغاز شود؟
این که آیا نوترکیبی به چهار رشته DNA نیازی دارد یا خیر، روشن نیست. همان طور که در بخش اول اشاره شد، نمی دانیم در آغازیانی که میوز تک مرحله ای رخ می دهد، کراسینگ اور رخ می دهد یا نه. دو تبیین برای طبیعت دو مرحله ای میوز وجود دارد که به علت پیچیدگی از اشاره به آن ها صرف نظر می کنیم.
آخرین نکته ای که درباره میوز ذکر می شود مهم است. فرایند نوترکیبی پیچیده است و نیاز دارد که دو رشته DNA به طور دقیق کنار هم به خط شوند، دقیقا در نقطه مشابهی بریده شوند و پس از تبادل در همان نقطه اتصال یابند. این فرایند همچنین به آنزیم نیاز دارد. نقصان در این سیستم آن را کاملا ناکارآمد می سازد. پس چگونه چنین سیستم پیچیده ای می تواند تکامل یابد؟
طبق معمول، پاسخ آن است که اجزای سیستم پیشتر تکامل یافته بوده اند تا عملکردی متفاوت ایفا کنند. آنزیم های مورد استفاده در نوترکیبی همان آنزیم های هستند که DNA آسیب دیده را ترمیم می کنند و به دقت بالا در برش و پیرایش مولکول DNA نیاز دارند. میوز مختص یوکاریوت هاست، اما بیشتر آنزیم های دخیل در میوز با عملکردی متفاوت در پروکاریوت ها حضور دارند.
گمان می کنیم نظریه ای که توصیف شد، توضیح قابل قبولی برای منشا میوز می باشد. سوالات بی پاسخی وجود دارند که ممکن است پاسخ آن ها را در مطالعه جانداران کنونی بیابیم. مخصوصا، نیاز داریم اطلاعات بیشتری درباره بوم شناسی جانداران در حال تناوب بین هاپلوئیدی و دیپلوئیدی و نیز ژنتیک آرکزوئر ها [3] به دست آوریم.
حال که با چگونگی ظهور میوز آشنا شدیم، زمان آن است که بدانیم انواع آمیزشی و تفاوت میان جنسیت ها چگونه حاصل شده اند. تقریبا همه آغازیان دو نوع آمیزشی + و - را دارند. گامت های نوع + تنها با گامت های نوع - لقاح می کنند و برعکس، گامت های نوع - نیز تنها با گامت های نوع + لقاح می کنند. نتیجه این است: گامت هایی که بسیار به هم شبیه هستند، با یکدیگر لقاح نخواهند داشت. اگر طبق نظریه پیشنهادی ما، مزیت لقاح، قدرت دورگه باشد، باید از لقاح گامت های مشابه پرهیز شود.
در نگاه نخست، این که چرا تنها دو نوع آمیزشی وجود دارد، جای سوال دارد. چرا انواع آمیزشی بیشتری با توان لقاح با یکدیگر وجود ندارد؟ به این ترتیب نسبت گامت هایی که قادرند با یک گامت خاص لقاح کنند، افزایش می یافت.
برای یک لحظه از موضوع منحرف شویم: چیزی شبیه این در گیاهان گلدار رخ می دهد. بیشتر آن ها هرمافرودیت بوده، تخم و گرده را با هم تولید می کنند اما خود ناسازگارند؛ بدین معنا که نمی توانند با خود لقاح داشته باشند. غالبا چندین نوع خود ناسازگار وجود دارد و گرده هر یک از آن ها می تواند با تمام انواع دیگر لقاح یابد.
پس چرا تنها دو نوع آمیزشی وجود دارد؟ یک پاسخ محتمل آن است که پیدایش دو نوع آمیزشی آسان تر از تعداد بیشتری بوده است، اما تکامل معمولا محدودیتی در تعداد نوآوری های خود ندارد.
امروزه به نظر می رسد تبیین این مسئله، میتوکندری ها و کلروپلاست ها باشند که در صورت وجودشان در یک جاندار، تنها از یکی از والدین به ارث می رسند. برای مثال، در جلبکی سبز موسوم به کلامیدوموناس (Chlamydomonas) که به خوبی مطالعه شده است، میتوکندری ها تنها از نوع - و کلروپلاست ها تنها از نوع + به ارث می رسند.
این قاعده توارث تک والدی برای اندامک های درون سلول، تقریبا جهانی است. در جانوران، میتوکندری از مادر به ارث می رسد. در گیاهان، کلروپلاست ها معمولا به تخمک منتقل می شوند، نه گرده. اما استثنا هایی وجود دارد: در مخروطیان، کلروپلاست ها به گرده منتقل می شوند.
چرا توارث تک والدی اندامک ها باید تا این حد جهان شمول باشد؟ پاسخ محتمل این است: اگر بگوییم میتوکندری ها از هر دو والد به ارث می رسند، عرصه برای تکامل اندامک های "خودخواه" فراهم می شود. در تقسیم سلولی، میتوکندری ها به طور تصادفی به دو سلول دختر منتقل می شوند. سیستمی برای تضمین انتقال نسخه ای از هر میتوکندری به همه سلول های دختر، مانند آن چه در مورد کروموزوم ها مشاهده می شود، وجود ندارد.
با توجه به این مطالب، اگر یک میتوکندری جهش یافته قادر به تکثیر سریع درون سلول باشد در حالی که چندان در تولید ATP موثر نباشد، در جمعیت سریعا منتشر می شود. ظهور چنین جهش یافته هایی، اتفاق می افتد اما به دلیل توارث تک والدی نمی توانند گسترش یابند و ممکن است سلولی را که در آن قرار دارند، از بین ببرند.
با توجه به لزوم توارث تک والدی اندامک ها، تکامل بیش از دو نوع آمیزشی دشوار است. در سال 1992، لارنس هرست و ویلیام هامیلتون مشاهدات کم نظیری ارائه کردند که نشان می داد این تبیین واقعا درست است.
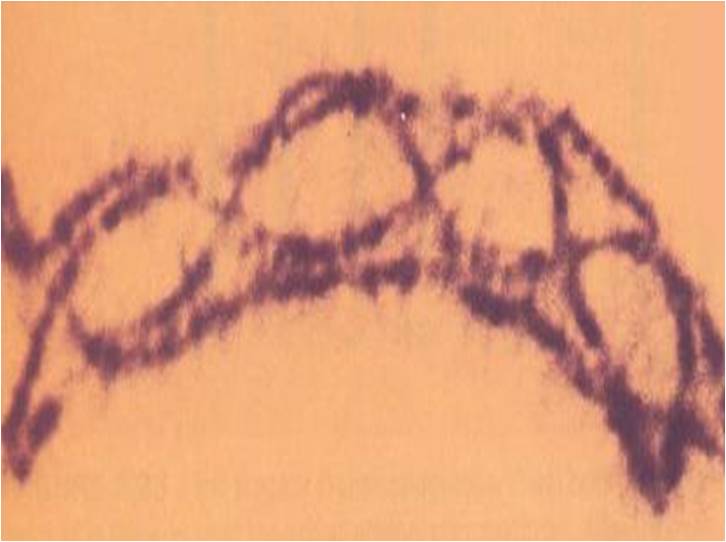
در آغازیان مژکدار همچون پارامسی، معمولا خبری از لقاح گامت ها نیست. به جای آن، دو سلول کنار یکدیگر قرار می گیرند یا اصطلاحا وارد هم یوغی می شوند. از هر یک هسته ای هاپلوئید به دیگری منتقل می شود بدون آن که سیتوپلاسم ها مخلوط شوند. پس از پایان هم یوغی، هر سلول دو مجموعه کروموزوم را از هر دو والد به ارث برده است اما هر یک تنها میتوکندری های خود را داراست. در این مژکداران همان طور که انتظار می رود، انواع آمیزشی ناسازگار فراوانی وجود دارد چرا که خطری برای گسترش اندامک های خودخواه وجود ندارد.
اما در گروهی از مژکداران به نام hypotrich ها، دو فرایند جنسی بدیل وجود دارد: هم یوغی و لقاح گامت ها. انواع آمیزشی متعددی برای هم یوغی وجود دارد اما تنها دو نوع از این انواع در لقاح نقش دارد. جالب است که این حقایق عجیب و حیرت انگیز در زمان تدوین این نظریه وجود نداشته اند اما در سایه این نظریه به خوبی معنا می یابند.
در جانوران و گیاهان، تقسیم وظیفه ای میان گامت متحرک و گامت حامل ذخایر غذایی لازم برای تکوین، وجود دارد. مدل هایی ریاضی وجود دارند که نشان می دهند اندازه فرد بالغ، متغیری است مرتبط با انیزوگامی [4]. مقایسه گونه های انیزوگامی و ایزوگامی نیز از این ارتباط حمایت می کند.

مژکداران hypotrich در حال هم یوغی، برگرفته از مدخل هم یوغی دانشنامه بریتانیکا
ولوکس (Volvox) سرده ای از جلبک های سبز است که خویشاوند کلامیدوموناس محسوب می شود اما کلنی های پرسلولی تشکیل می دهد. کلنی این جانداران عبارت است از کره ای تو خالی از سلول های سبز مژکدار. در گونه هایی که کلنی کوچک است، گامت ها همگی متحرک و هم اندازه هستند [ایزوگامی]. در کلنی های دارای اندازه متوسط، گامت ها متحرک هستند اما دارای اندازه متفاوتی هستند. گونه های واجد بزرگترین کلنی، دارای گامت های بزرگ غیر متحرک و گامت های کوچک متحرک هستند [انیزوگامی].
به کمک ولوکس، می توانیم در مقیاسی کوچک، آن مسیر تکاملی را که نیاکان جانوران و گیاهان طی کرده اند مشاهده کنیم. جنس نر [گامت متحرک با اندازه کوچک تر]، نخستین جنسیتی است که تکامل یافته و جنس ماده [گامت غیر متحرک و بزرگتر] دومین جنسیت. احتمالا به دنبال انیزوگامی، صفات ثانویه جنسی تکامل یافته اند. علت این امر، آن است که تولید سلول تخم هزینه بر است و تولید اسپرم، کم هزینه. بنابراین نر ها در آمیزش های مکرر، از ماده ها مفیدتر هستند.
عدم تقارن سلول های جنسی نر و ماده، غالبا موجب تفاوت در شکل و اندازه میان جنسیت ها شده است. این خلاصه ای شدیدا ساده از موضوعی پیچیده است. به طور خلاصه می توان تاریخ تفکیک جنسیت ها را به سه مرحله تقسیم کرد: اول، تکامل انواع آمیزشی + و - به دلیل نیاز به توارث تک والدی. دوم، تکامل جنس نر و ماده با تولید گامت های متحرک و ذخیره کننده مواد غذایی. سوم، تکامل صفات ثانویه جنسی در برخی دودمانه ها که بخشی از آن در نتیجه رقابت برای آمیزش و بخش دیگر حین تقسیم وظایف مراقبت از فرزندان، ظاهر شده است.
متن فوق برگرفته از کانال آموزش علم می باشد که گزیده ای از کتاب The Origins of Life: from the birth of life to the origins of language را در بر دارد.
پانویس ها:
1- عبارت لاین درون آمیخته (inbred line) یا سویه درون آمیخته (inbred strain) به افرادی از یک گونه اطلاق می شود که به علت درون آمیزی طولانی مدت بسیار به یکدیگر شباهت دارند.
2- این پدیده انتقال عرضی ژن (horizontal gene transfer) نامیده می شود و مختص باکتری ها نیست.
3- آرکزوئر ها (Archezoa) فرمانرویی است که آغازیان فاقد میتوکندری را شامل می شود.
4- اگر در تولید مثل جنسی، گامت ها از نظر شکل و اندازه با یکدیگر متفاوت باشند، چنین حالتی را انیزوگامی (Anisogamy) می نامند. حالت مقابل انیزوگامی، ایزوگامی (Isogamy) نام دارد.